栄養学科 > 栄養学の歴史 > 総目次 > 次の実験
15: ビタミン前駆体としてのトリプトファンの役割
The Journal of Nutrition Vol. 127 No. 5 May 1997, pp. 1017S-1053S
Paper 15: Tryptophan's Role as a Vitamin Precursor (Krehl et al., 1945)
LaVell M. Henderson, Professor Emeritus, Department of Biochemistry, University of Minnesota, St. Paul, MN
リンク:栄養学の考え方を変えさせた実験 (原論文)
イヌの黒舌病がペラグラに対応することが認識されると、1925年ごろ実験栄養学者たちはイヌをペラグラ病因の研究に使うようになった。カサール(Casal)がこの症状群を記載してから二百年後に、新しいB群ビタミンであるナイアシンの欠乏によることが、エルビーエムらの実験によって示された(Elvehjem et al.1938)。
L. J. Teplyは種々の食物のナイアシン活性をSnell and Wrightの微生物法で測定した。予想に反して、トウモロコシには黒舌病とペラグラを予防するのに充分なナイアシンを含んでいた。クレールと協同研究者たちはこのことを確認したが、ショ糖-カゼイン食餌のときに比べてトウモロコシ食餌のときにはイヌのナイアシン要求量は3倍であることを見いだした。ショ糖-カゼイン食餌で飼ったラットは食餌にナイアシンを添加する必要が無かったので、トウモロコシ食餌で飼ったラットで試してみた。Krehl et al. 1945aは、15%カゼインの40%をひき割りトウモロコシ粉 (corn grits) で置き換えたところ、成長は1週あたり25gであったのが4gになった、と報告した。ナイアシンを加えると成長が元に戻ったので、この条件でラットはナイアシンを必要とすることが判った。トウモロコシは必須アミノ酸のレベルが異常に低い穀物である。2月後にKrehl et al. (1945b)は、100g食餌あたり40mgのトリプトファンがナイアシンの代わりとしてひき割りトウモロコシ粉による成長抑制を元に戻す、と報告した(表 1)。
| ||||||||||||||||||||||||||||||||||
食餌に1mg%必要なナイアシンとモル比で40倍のトリプトファンの互換性のもっとも妥当な説明は、トリプトファンがナイアシンに転換することであった。ナイアシン欠乏において40%トウモロコシ粉の代わりを2%の酸加水分解カゼイン(トリプトファンは破壊)がすることは、他のアミノ酸が増えるとナイアシン欠乏を誘導する可能性を示唆した。この効果は”アミノ酸インバランス”と名付けられた。ナイアシン欠乏をもっとも起こしやすいのはトレオニンとリシンであり(Hankes et al. 1948)、この効果を最大にするには硫黄アミノ酸の追加が必要であった。したがって、その後の実験では基本食餌に0.2%のL-シスチンが加えられた(表1)。9% カゼイン + 0.2% L-シスチンで飼ったラットにたいする他のアミノ酸の影響は次の図によって説明される。
トレオニンのような必須アミノ酸が最適レベルより少し低い程度に供給されると、適度の成長が起きて、ぎりぎりの量のトリプトファンがタンパク合成とナイアシン合成の両方を満足させます。食事中のトレオニン量が増えるとタンパク質合成が増え、トリプトファンは別の経路であるナイアシン合成を犠牲にして、タンパク質合成の方に引っ張られてしまい、ビタミン欠乏が起きる。トレオニンの効果についてのこの説明は、リシン、バリン、ロイシン、イソロイシンが成長の阻害を起こしトリプトファンまたはナイアシンで逆転できることにより、支持される(Koeppe and Henderson 1955)。
ラットについての多くの同位体標識実験はトリプトファンのナイアシンへの変換を確認した。この反応についての最初の事実はアカパンカビ(Neurospora crassa)変異種の実験で得られた(図 1)。この図の最初の4つの化合物は動物の系で認められ、キノリン酸は哺乳類でトリプトファンからの排泄物質として同定され、ラットおよびナイアシン非生産性変異株アカパンカビであまりナイアシンの代わりにならないものであった(Henderson 1949)。
 図 1 アカパンカビおよびラットにおいてトリプトファンがナイアシンになる経路の中間代謝産物
図 1 アカパンカビおよびラットにおいてトリプトファンがナイアシンになる経路の中間代謝産物
3-ヒドロキシアントラニル酸を哺乳類肝臓とインキュベートすると、キノリン酸とピコリン酸が生成した (Fig. 2)。両方ともin vivoでも二酸化炭素にまで分解されない。これらがトリプトファンからグルタール酸への分解過程の中間物であることはGholson et al. (1962)の報告によって確立された。ピコリン酸とキノリン酸はトリプトファン分解の中間産物が環状のシッフ塩基(訳注:1級アミンとカルボニル基の縮合産物)を作ることによって出来ることは明らかであり、トリプトファン分解における副産物とみなすことができる。
外因性のキノリン酸が食餌でナイアシンの代用になり難いことから、ナイアシン生成におけるキノリン酸の役割に疑いがもたれた。この制限はたぶんキノリン酸塩が細胞に入れないことによるのであろう。キノリン酸は強酸であり非解離形で細胞内に入らなければならないので、膜を通過するのに適当なpHは生理的pHよりずっと低いからである。ナイアシン生成におけるキノリン酸の役割は西塚と早石(Nishizuka and Hayaishi 1963)の研究によって解明された。彼らはラット肝臓の可溶性酵素によってキノリン酸が5-ホスホリボシル1-ピロリン酸の存在のもとでニコチン酸モノヌクレオチドになることを示した (図2)。
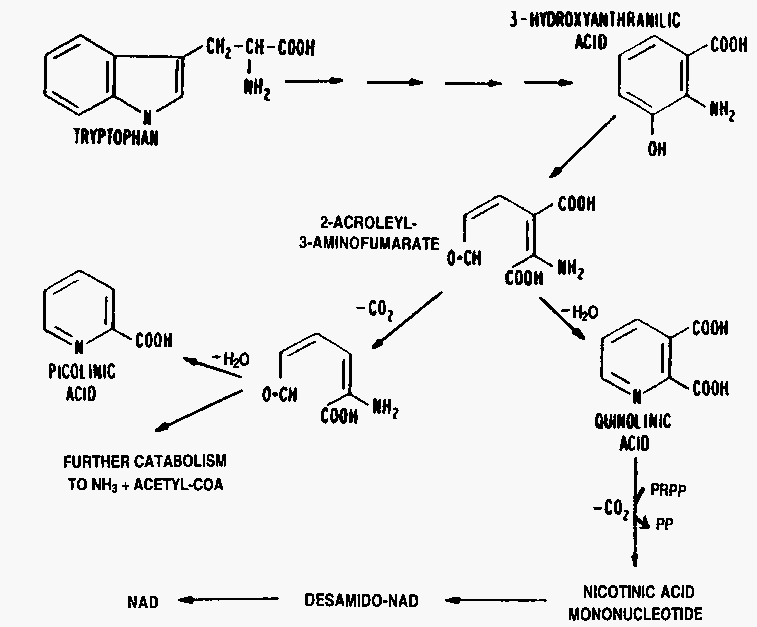 図 2 トリプトファンの全代謝およびナイアシンを含むピリジニウム化合物への変化の簡略スキーム
図 2 トリプトファンの全代謝およびナイアシンを含むピリジニウム化合物への変化の簡略スキーム
トリプトファンがナイアシンになる効率が動物種によって大きく異なるのは2-アクロレイル 3-アミノフマール酸が脱炭酸される割合によるようである(図2)。ネコの肝臓では86%が脱炭酸され、9%だけがキノリン酸になる (Suhadolnik et al. 1957)。反対の割合の例をあげると、ラット肝臓は12%だけを脱炭酸し、残る80%はキノリン酸になる。イヌやヒトを含む他の8種の肝臓はこの両極端の中央に位置する。ネコではトリプトファンがナイアシンの代わりにならないが、ラットはふつうのレベルのトリプトファン摂取ではナイアシンを必要としない。ヒトとイヌはトリプトファンがナイアシンの代わりをする能力には限りがある。したがって、適当にトリプトファンを摂取していても臨床的な欠乏が起きる。
南ヨーロッパのトウモロコシを食料とする貧しい人々でペラグラが流行し、西半球の土着の人たちに見られないことは、アメリカ土着の人たちはコーンミールからトルティージャ(tortilla)を作るときに石灰水などアルカリ性液を使うことによって、説明されてきた。Krehl et al. (1945b)の50年前の観察はナイアシン必要量が種によって違うことを説明し、トウモロコシにトリプトファンが非常に少ない事実から、トウモロコシの消費とペラグラの関係を部分的に説明してきた。この関係を完全に理解するには、アルカリと一緒に熱するとナイアシンになる仮説的物質の同定が必要である。
文献
Elvehjem C. A., Madden R. J., Strong F. M., Woolley D. W. The isolation and identification of the anti-black tongue factor. J. Biol. Chem. 1938; 123:137-149
Gholson, R. K., Nishizuka, Y., Ichiyama, A., Kawai, H., Nakamura, S. & Hayaishi, O. (1962) New intermediates in the catabolism of tryptophan in mammalian liver. J. Biol. Chem. 237: PC2043-2045.
Hankes L. V., Henderson L. M., Brickson W. L., Evlehjem C. A. Effect of amino acids on the growth of rats on niacin-tryptophan deficient rations. J. Biol. Chem. 1948; 174:873-881
Henderson L. M. Quinolinic acid metabolism II. Replacement of nicotinic acid for the growth of the rat and Neurospora. J. Biol. Chem. 1949; 181:677-685
Koeppe O. J., Henderson L. M. Niacin-tryptophan deficiency resulting from imbalances in amino acid diets. J. Nutr. 1955; 55:23-33
Krehl, W. A., Teply, L. J. & Elvehjem, C. A. (1945a) Corn as an etiological factor in the production of nicotinic acid deficiency in the rat. Science 101: 283.
Krehl W. A., Teply L. J., Sarma P. S., Elvehjem C. A. Growth-retarding effect of corn in nicotinic acid-low rations and its counteraction by tryptophane. Science 1945b; 101:489-490
Nishizuka, Y. & Hayaishi, O. (1963) Enzymic synthesis of niacin nucleotides from 3-hydroxyanthranilic acid in mammalian liver. J. Biol. Chem. 238: PC483-485.
Suhadolnik R. J., Stevens C. O., Decker R. H., Henderson L. M., Hankes L. V. Species variation in metabolism of 3-hydroyanthranilate to pyridinecarboxylic acids. J. Biol. Chem. 1957; 228:973-982
Copyright(C) 2006-2008 Department of Nutritional Sciences. All Rights Reserved.