栄養学科 > 栄養学の歴史 > 総目次 > 次の実験
14: 身体構成要素の動的状態
The Journal of Nutrition Vol. 127 No. 5 May 1997, pp. 1017S-1053S
Paper 14: The Dynamic State of Body Constituents (Schoenheimer, 1939)
Robert E. Olson, Department of Pediatrics, College of Medicine, University of South Florida, Tampa, FL
リンク:栄養学の考え方を変えさせた実験 (原論文)
コロンビア大学におけるシェーンハイマー (Rudolf Schoenheimer) と彼の共同研究者は1933年から1941年の間に脂肪酸およびアミノ酸の代謝について我々の理解に革命的変化をもたらし、代謝回転の概念にみちびいた。
シェーンハイマーは1898年にベルリンに生まれ、1922年に医学の学位をベルリン大学より受けた。1年間の臨床修練の後でライプチッヒのKarl Thomas (生化学者)のもとで1923年から1926年までポストドック研究を行い、1926年から1933年までフライブルクのLudwig Aschoff (著名な病理学者)のもとでコレステロール代謝の研究を行った。1933年のドイツ政変により彼はアメリカに亡命してコロンビア大学生化学講座で研究することになった。重水素を1932年に発見したHarold UreyおよびUreyの学生でコロンビアの生化学科に移っていたリッテンバーグ (David Rittenberg)と、ここで彼は知り合いになった。シェーンハイマーとリッテンバーグは一緒に重水や重水素と15Nで標識したアミノ酸などの種々の同位体を用いる実験を計画した。これらの実験は脂肪とタンパク質の代謝の概念に革命を起こした。
シェーンハイマーが中間代謝の同位元素研究をはじみた時には、体脂肪やタンパク質など身体の主な成分は化学的に安定であると、一般に考えられていた。”すり切れ(wear and tear)”による組織の置き換え以外に、食餌と体内の種々の臓器の間には栄養素分子の交換は無いとされていた。体の貯蔵脂肪はエネルギーの予備であって、食事制限のときのみ招集されるものであり、そうでないときには、脂肪代謝は食餌中の脂肪を酸化してエネルギーを作るときにのみ起きるものであった。
タンパク質についても同様な見解が、主としてドイツのルーブナー(Rubner)やアメリカのフォリン(Folin)の研究によって支持された。1905年にフォリンは尿中窒素化合物の排泄の研究をもとにして、窒素代謝は内因性(組織)と外因性(食餌)の2相に分けられるとした (Folin 1905)。マッカラム(E. V. McCollum et al. 1939)は”the Newer Knowledge of Nutrition"の5版でフォリンを賞賛して書いた。”フォリンはタンパク質代謝の問題を主流から解決する天才を持っていて、彼の解釈にほとんど誰も反対しない”、と。フォリンの見解はタンパク質代謝のパラダイムとなり、シェーンハイマーが研究を始めた1930年代の半ばまで続いた。
脂肪代謝の研究
身体構成成分の代謝回転の最初の研究として、シェーンハイマーは体脂肪合成の追求に重水を使用した。水のプロトンはNADPHによる還元過程で脂肪酸の合成に取り入れられた。図 1は19日の間における貯蔵脂肪中の脂肪酸への濃縮を示す。マウス全脂肪酸の重水素量は6日で最大に達し、ハーフタイムは2.5日であった。濃縮全体から計算すると、すべての脂肪酸(主として脂肪組織)の14%がこの実験で置き換えられた。炭水化物が豊富な食餌で飼ったマウスにおける脂肪酸の分解を示すために、同体重のマウスを重水素標識の脂肪を20%、全粒パン80%からなる食餌で5日のあいだ飼育した。マウスは間隔を置いて殺した。標識脂肪は最初の合成試験と同じ速度で分解した (図 1)。シェーンハイマーは[2H]-パルミチン酸がラットの脂肪に蓄積しただけでなく、ステアリン酸やオレイン酸をふくむ他の脂肪酸に変換することを示した。しかし、すでに必須脂肪酸であることが示されていたリノール酸には変換しなかった。
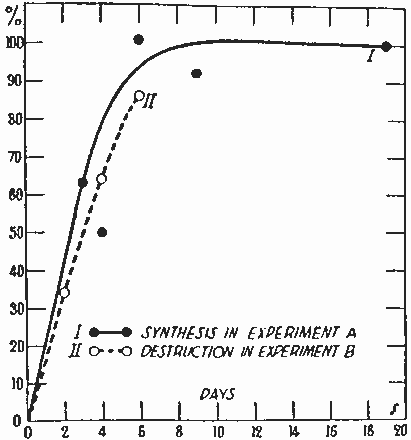 図 1. マウスにおける脂肪酸の合成と分解 (Schoenheimer and Rittenberg 1936)。 Journal of Biological Chemistryより転載許可
図 1. マウスにおける脂肪酸の合成と分解 (Schoenheimer and Rittenberg 1936)。 Journal of Biological Chemistryより転載許可
タンパク質代謝の研究
シェーンハイマーと共同研究者たちは次にタンパク質代謝の研究に転じた。彼らは水素と窒素で二重標識をしたL(-)ロイシンを合成した。これは3.60 atoms % excess (APE) の重水素と6.54APEの15Nを含んでいた。(訳注:たとえば窒素については、100原子の14N + 15Nのうちの15N原子数がatoms %(At%)であり、バックグラウンドのAt%を引いたのがAPEである。)この両方の同位体濃度は充分に高く、数百倍のふつうのロイシンと混ぜても質量分析法で測定できた。標識ロイシンを混ぜた餌を成熟ラットに与えて3日のあいだ観察した。この間に体重の変化は無かった。3日目の終わりに排泄物および組織を分析した。摂取した15Nの約30%は排泄物(糞中に2.2%、尿中に27%)に見つかった。体タンパク質に残ったのは65%の活性であり、同位元素Nは主として組織タンパク質に組み入れられたことが示された。この発見は外因性タンパク質代謝についてのフォリンの見解と矛盾した。
食餌窒素の組み入れは臓器が違うと同じでないことが示された。表 1に示されるように、内臓、血清、腸管はもっとも活性が高かった。筋タンパク質の活性は高くなかったが、動物で筋の量はもっとも高いので、15Nの濃度は低くても同位体の絶対量は高かった。事実、摂取した同位体の66%は筋に回収され、33%だけが内臓全体に回収された。
| ||||||||||||||||||||
ロイシンは種々の臓器や組織に組み入れられただけでなく、表 2に示すようにリシンを例外として15Nは他の組織アミノ酸に出現した。この交換反応でとくに顕著なのはグルタミン酸とアスパラギン酸であった。シェーンハイマーが観察したこのアミノ基転位は1937年に酵素学的にBraunsteinとKretzmann (訳注:ソヴィエトの生化学者)によって説明された。アルギニンの15Nは腎臓よりも肝臓に濃縮された。これは主としてKrebsとHenseleitが1932年に発見した肝臓のオルニチン-尿素サイクルによるものである。
| |||||||||||||||||||||||||||||||||
摂取した二重標識ロイシンの両方の同位体量を測定すると、D/15Nは、摂取時には100:182であったが、屍体では100:108であった。このことは摂取ロイシンの標識窒素の1/3以上がふつうの窒素で置き換えられたことを示していた。このことはアミノ基転位がアミノ酸の重要な反応であることの証明であった。
シェーンハイマーと共同研究者たちはタンパク質の少ない食餌で飼った未成熟ラットに[15N]クエン酸アンモニウムを与えた。この自然ではない食餌によってラットは体重が減少したが、しかし体タンパク質の急速な消失にもかかわらず、食餌のアンモニアから新しいアミノ酸が合成された。グルタミンとアスパラギンのアミド Nは、グルタミン酸のα-アミノ基と同じように15Nが濃縮された。グルタミン酸合成酵素、グルタミン酸デヒドロゲナーゼ、カルバミル・リン酸合成酵素はアミノ酸へアンモニアを固定する重要な酵素として知られている。アルギニン内の大部分の同位体はアミヂン基にありアルギナーゼで除かれ、オルニチン-尿素サイクルによると、オルニチンには非常に少量しか残らない。
これらの実験で [15N]アンモニアはクレアチンに組み入れられた。これはアルギニンがクレアチンの前駆体であることを示したBorsook and Dubnoffの1941年の研究から想定できた。シェーンハイマーが示したのは[15N]クレアチンからのクレアチニンの生成は29日の間に同位体希釈が起きない自発反応であることであった。クレアチニン生成曲線は身体全体のクレアチンの2%が毎日尿クレアチニンになる(新合成によって置き換えられる)ことであった。このことはまた尿中クレアチニン排泄が一定であることを説明した。これはタンパク質の"外因性代謝" を示すものとしてフォリンが説明していたものであった。
シェーンハイマーたちは、前に論じたフォリンの仮説に正式に反対して次ぎのように述べた:”我々の発見は窒素に2つの型の区別を必要とするどのような理論とも両立することはできない.....排泄された窒素は、食餌性窒素とかなり大量の反応性組織窒素の相互作用に始まる代謝プールの一部と考えるべきであろう” (Schoenheimer et al. 1939)。
要約すると、シェーンハイマーは脂肪とタンパク質代謝について新しい動的なパラダイムを導入して、フォリンその他の静的な観点を置き換えた。事実、主要な身体構成成分が動的な生化学システムであるという考えは、最近になり代謝のすべての様相に拡張されてきている。このシステムには物質輸送、造血、内分泌、サイトカイン活性、さらにはゲノムまでが含まれる。
文献
Borsook H., Dubnoff J. W. The formation of glycocyamine in animal tissues. J. Biol. Chem. 1941; 138:389-394
Braunstein A. E., Kritzmann M. G. Über den Ab- und Aufbau von Aminosäuren durch Umaminierung. Enzymologia 1937; 2:129-140
Folin O. A theory of protein metabolism. Am. J. Physiol. 1905; 13:117-138
Krebs H. A., Henseleit K. Untersuchungen über die Harnstoffbildung im Tierkörper. Z. Physiol. Chem. 1932; 210:33-45
McCollum, E. V., Orent-Keiles, E. & Day, H. G. (1939) The Newer Knowledge of Nutrition, 5th ed., pp. 80-82. MacMillan, New York, NY.
Schoenheimer, R. (1942) The Dynamic State of Body Constituents. Hafner Publishing, New York, NY.
Schoenheimer R., Ratner S., Rittenberg D. Studies in protein metabolism. X. The metabolic activity of body proteins investigated with L(-)leucine containing two isotopes. J. Biol. Chem. 1939; 130:703-732
Schoenheimer R., Rittenberg D. Deuterium as an indicator in the study of intermediary metabolism, VI. Synthesis and destruction of fatty acids in the organism. J. Biol. Chem. 1936; 114:381-396
Copyright(C) 2006-2008 Department of Nutritional Sciences. All Rights Reserved.